|
||
|
立体选择性羰基还原酶及其在手性醇合成中的应用
化工进展
2021, 40 (3):
1142-1160.
DOI:10.16085/j.issn.1000-6613.2020-2124
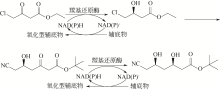
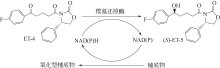
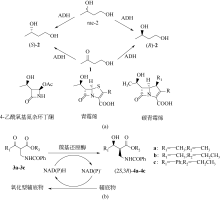
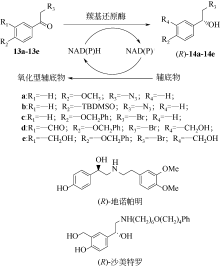
手性醇是重要的医药中间体与精细化工品,立体选择性羰基还原酶催化制备手性醇具有重要的研究与应用价值,受到国内外科学家和工程师的高度关注。本文主要围绕羰基还原酶的发现与分类、催化羰基还原反应的活性与立体选择性机制、酶的筛选挖掘与分子改造技术、辅酶还原型烟酰胺腺嘌呤二核苷酸(磷酸)的再生方法、羰基还原酶催化合成手性醇医药中间体与精细化学品技术开发与应用等方面展开综述。重点阐述了羰基还原酶催化制备降血脂、抗细菌或病毒感染、抗肿瘤、抗抑郁症、抗癫痫等重要疾病治疗药物中间体及脂肪族、芳香族手性醇精细化工品的国内外技术进展与应用,为高效能立体选择性生物催化剂的创制和手性化合物的生物合成提供理论借鉴和成功范例。 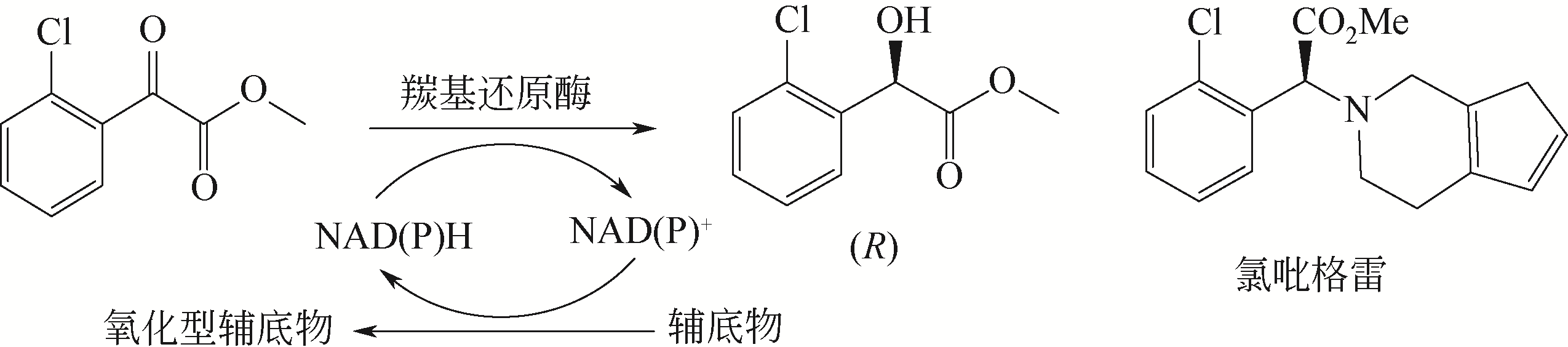 View image in article
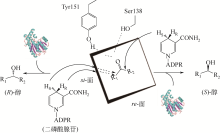
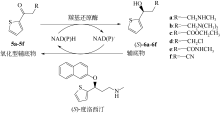
图12
(S)-氯吡格雷手性中间体的酶法合成
正文中引用本图/表的段落
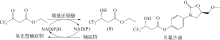
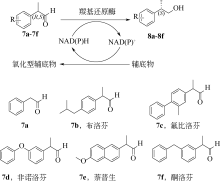
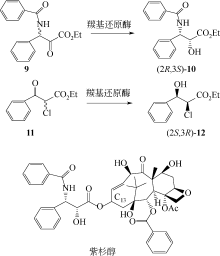
(S)-氯吡格雷(Plavix)是一种选择性血小板凝集抑制剂,由法国赛诺菲(Snaofi)公司开发,用于治疗因血栓引起的心肌梗死和缺血性卒中,是目前世界上最畅销的药物之一。在已报道的(S)-氯吡格雷合成途径中,(R)-邻氯扁桃酸甲酯的合成是关键步骤(图12)。Jeong等[114]利用邻氯苯乙酮酸甲酯为底物,以Saccharomyces cerevisiae全细胞为催化剂,不对称还原获得(R)-邻氯扁桃酸甲酯,e.e.达96.1%。一株来源于Thermus thermophilus的羰基还原酶也得到纯化与鉴定,使用该酶催化邻氯苯乙酮酸甲酯的还原,产率达95%,产物e.e.为92%[115]。Ma等[116]对C. glabrata基因组进行分析,挖掘获得一株新型羰基还原酶CgKR1,在与葡萄糖脱氢酶共表达构建辅酶再生体系后,催化邻氯苯乙酮酸甲酯不对称还原生成(R)-邻氯扁桃酸甲酯,底物浓度达300g/L,产率为87%,e.e.达98.7%。Ni等[117]将来源于Bacillus sp.的醛酮还原酶与葡萄糖脱氢酶共表达后,催化(R)-邻氯扁桃酸甲酯的合成,底物 浓度进一步提升至500g/L,时空产率达812g/(L·d),e.e.>99%。Xu等[39]发现,甘油可提升NADPH依赖型醛酮还原酶YtbE的热稳定性与操作稳定性,在1mol/L底物浓度下,使(R)-邻氯扁桃酸甲酯的产率由70.5%提高到96.6%。Zheng等[118]对羰基还原酶CgKR1 进行分子改造,获得CgKR1-F92C/F94W,用于系列手性醇医药中间体的制备,其中催化(R)-邻氯扁桃酸甲酯合成的活性提高了8倍。
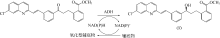
(S)-氯吡格雷(Plavix)是一种选择性血小板凝集抑制剂,由法国赛诺菲(Snaofi)公司开发,用于治疗因血栓引起的心肌梗死和缺血性卒中,是目前世界上最畅销的药物之一.在已报道的(S)-氯吡格雷合成途径中,(R)-邻氯扁桃酸甲酯的合成是关键步骤( A tailor-made self-sufficient whole-cell biocatalyst enables scalable enantioselective synthesis of (R)-3-quinuclidinol in a high space-time yield 1 2019 ... (R)-(-)-3-奎宁醇含有一个含氮双环,是抗哮喘药物他沙利定、瑞伐托酯等药物合成的重要前体.不同的(R)-(-)-3-奎宁醇化学制备方法包括外消旋3-奎宁醇的动力学拆分、3-奎宁酮不对称还原等都已获报道[ Proposed correlation of data for isothermal two-phase, two-component flow in pipes 1 1949 ... 立体选择性羰基还原酶(carbonyl reductase, EC 1.1.1.148)可在还原型辅酶烟酰胺腺嘌呤二核苷酸(磷酸)[NAD(P)H]存在的情况下,催化手性酮不对称还原得到手性醇[ Biocatalytic ketone reduction: a green and efficient access to enantiopure alcohols 2 2012 ... 立体选择性羰基还原酶(carbonyl reductase, EC 1.1.1.148)可在还原型辅酶烟酰胺腺嘌呤二核苷酸(磷酸)[NAD(P)H]存在的情况下,催化手性酮不对称还原得到手性醇[
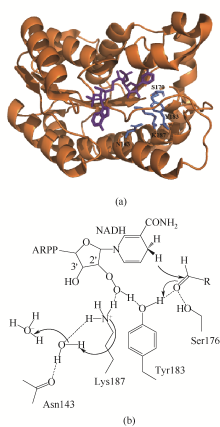
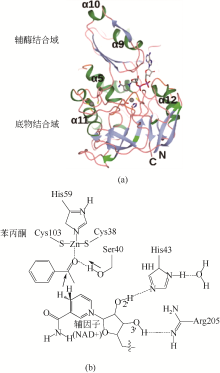
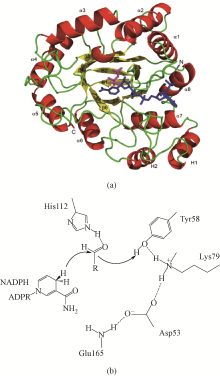
醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[ Subdivision of the MDR superfamily of medium chain dehydrogenases/reductases through iterative hidden Markov model refinement 1 2010 ... 中链脱氢酶(MDRs)是一类约含350个氨基酸残基的锌或非锌-依赖型脱氢酶.根据迭代隐马尔可夫模型(HMMs),中链脱氢酶可分为86类,包括MDR001-ADH、MDR002-PTGR、MDR003-FAS及MDR010-CAD等,不同种类间序列同源性在40%~90%之间.中链脱氢酶一般是以单聚体、二聚体或四聚体的形式存在.每个亚基含有两个结构部分[ Superfamilies SDR and MDR: from early ancestry to present forms. emergence of three lines, a Zn-metalloenzyme, and distinct variabilities 1 2010 ... 中链脱氢酶(MDRs)是一类约含350个氨基酸残基的锌或非锌-依赖型脱氢酶.根据迭代隐马尔可夫模型(HMMs),中链脱氢酶可分为86类,包括MDR001-ADH、MDR002-PTGR、MDR003-FAS及MDR010-CAD等,不同种类间序列同源性在40%~90%之间.中链脱氢酶一般是以单聚体、二聚体或四聚体的形式存在.每个亚基含有两个结构部分[ The aldo-keto reductase superfamily homepage 1 2003 ... 醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[ The diversity of microbial aldo/keto reductases from Escherichia coli K12 1 2013 ... 醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[ Comparative anatomy of the aldo-keto reductase superfamily 1 1997 ... 醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[ Aldo-keto reductase (AKR) superfamily: genomics and annotation 1 2009 ... 醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[ Human aldo-keto reductases: function, gene regulation, and single nucleotide polymorphisms 1 2007 ... 醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[ Biocatalytic properties of a recombinant aldo-keto reductase with broad substrate spectrum and excellent stereoselectivity 2011 Characterization and identification of three novel aldo-keto reductases from Lodderomyces elongisporus for reducing ethyl 4-chloroacetoacetate 2014 Genomic mining-based identification of novel stereospecific aldo-keto reductases toolbox from Candida parapsilosis for highly enantioselective reduction of carbonyl compounds 2014 Improved o-chlorobenzoylformate bioreduction by stabilizing aldo-keto reductase YtbE with additives 1 2014 ... (S)-氯吡格雷(Plavix)是一种选择性血小板凝集抑制剂,由法国赛诺菲(Snaofi)公司开发,用于治疗因血栓引起的心肌梗死和缺血性卒中,是目前世界上最畅销的药物之一.在已报道的(S)-氯吡格雷合成途径中,(R)-邻氯扁桃酸甲酯的合成是关键步骤( Microbial aldo-keto reductases 2 2002 ... 醛酮还原酶(AKRs)是一类含有320个左右氨基酸的非金属依赖型羰基还原酶,广泛存在于动物、植物、原核生物中,并大多以单体形式存在[
本文的其它图/表
|

{kind=link}