Multiscale evolutionary dynamics of host-associated microbiomes
1
2018
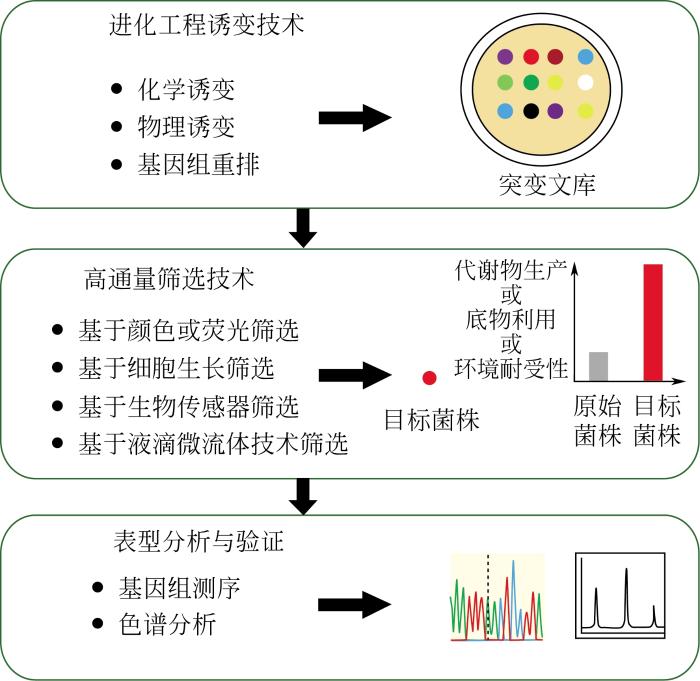
... 过去几十年,菌种进化工程研究和应用取得了很多突破性的进展,研究人员开发了一系列物理、化学诱变技术实现了不同基因尺度的微生物进化[1](如单核苷酸突变、小片段的插入和缺失、基因组重排)能够在较短的时间内产生大量突变体,随后通过合适的筛选方法可以快速从突变文库中筛选出理想的突变菌株.作为一种模拟自然界进化的方法,菌种进化工程弥补了传统理性设计的不足,它不依赖复杂的微生物生理学和信息学知识的指导,而是通过随机诱变的方式获得某种特性显著提高的突变体,同时随着相关组学技术的发展,研究人员能够快速从优良的突变体中分析获得有用的突变信息,并为理性设计提供新的指导[2].基于这些技术优势,菌种进化工程被广泛应用于微生物代谢工程领域,以提高微生物的多种生理和生产特性,如环境耐受性[3]、底物利用率[4]以及代谢物生产[5]等. ...
Metabolic engineering: past and future
1
2013
... 过去几十年,菌种进化工程研究和应用取得了很多突破性的进展,研究人员开发了一系列物理、化学诱变技术实现了不同基因尺度的微生物进化[1](如单核苷酸突变、小片段的插入和缺失、基因组重排)能够在较短的时间内产生大量突变体,随后通过合适的筛选方法可以快速从突变文库中筛选出理想的突变菌株.作为一种模拟自然界进化的方法,菌种进化工程弥补了传统理性设计的不足,它不依赖复杂的微生物生理学和信息学知识的指导,而是通过随机诱变的方式获得某种特性显著提高的突变体,同时随着相关组学技术的发展,研究人员能够快速从优良的突变体中分析获得有用的突变信息,并为理性设计提供新的指导[2].基于这些技术优势,菌种进化工程被广泛应用于微生物代谢工程领域,以提高微生物的多种生理和生产特性,如环境耐受性[3]、底物利用率[4]以及代谢物生产[5]等. ...
In vivo evolutionary engineering for ethanol-tolerance of Saccharomyces cerevisiae haploid cells triggers diploidization
1
2017
... 过去几十年,菌种进化工程研究和应用取得了很多突破性的进展,研究人员开发了一系列物理、化学诱变技术实现了不同基因尺度的微生物进化[1](如单核苷酸突变、小片段的插入和缺失、基因组重排)能够在较短的时间内产生大量突变体,随后通过合适的筛选方法可以快速从突变文库中筛选出理想的突变菌株.作为一种模拟自然界进化的方法,菌种进化工程弥补了传统理性设计的不足,它不依赖复杂的微生物生理学和信息学知识的指导,而是通过随机诱变的方式获得某种特性显著提高的突变体,同时随着相关组学技术的发展,研究人员能够快速从优良的突变体中分析获得有用的突变信息,并为理性设计提供新的指导[2].基于这些技术优势,菌种进化工程被广泛应用于微生物代谢工程领域,以提高微生物的多种生理和生产特性,如环境耐受性[3]、底物利用率[4]以及代谢物生产[5]等. ...
Comparative transcriptomes reveal novel evolutionary strategies adopted by Saccharomyces cerevisiae with improved xylose utilization capability
1
2017
... 过去几十年,菌种进化工程研究和应用取得了很多突破性的进展,研究人员开发了一系列物理、化学诱变技术实现了不同基因尺度的微生物进化[1](如单核苷酸突变、小片段的插入和缺失、基因组重排)能够在较短的时间内产生大量突变体,随后通过合适的筛选方法可以快速从突变文库中筛选出理想的突变菌株.作为一种模拟自然界进化的方法,菌种进化工程弥补了传统理性设计的不足,它不依赖复杂的微生物生理学和信息学知识的指导,而是通过随机诱变的方式获得某种特性显著提高的突变体,同时随着相关组学技术的发展,研究人员能够快速从优良的突变体中分析获得有用的突变信息,并为理性设计提供新的指导[2].基于这些技术优势,菌种进化工程被广泛应用于微生物代谢工程领域,以提高微生物的多种生理和生产特性,如环境耐受性[3]、底物利用率[4]以及代谢物生产[5]等. ...
Selecting the best: evolutionary engineering of chemical production in microbes
1
2018
... 过去几十年,菌种进化工程研究和应用取得了很多突破性的进展,研究人员开发了一系列物理、化学诱变技术实现了不同基因尺度的微生物进化[1](如单核苷酸突变、小片段的插入和缺失、基因组重排)能够在较短的时间内产生大量突变体,随后通过合适的筛选方法可以快速从突变文库中筛选出理想的突变菌株.作为一种模拟自然界进化的方法,菌种进化工程弥补了传统理性设计的不足,它不依赖复杂的微生物生理学和信息学知识的指导,而是通过随机诱变的方式获得某种特性显著提高的突变体,同时随着相关组学技术的发展,研究人员能够快速从优良的突变体中分析获得有用的突变信息,并为理性设计提供新的指导[2].基于这些技术优势,菌种进化工程被广泛应用于微生物代谢工程领域,以提高微生物的多种生理和生产特性,如环境耐受性[3]、底物利用率[4]以及代谢物生产[5]等. ...
Bacterial microcolonies in gel beads for high-throughput screening of libraries in synthetic biology
1
2017
... 菌种进化工程成功的关键是获得多样性的突变文库和开发高通量的筛选方法(图1).突变文库越大,突变体数量越多,则筛选到目标表型菌株的可能性就会越大.而对于庞大的突变体文库,高通量的筛选方法的开发是至关重要的[6],在一项研究中,Liu等[7]利用液滴微流控技术在短短的13h内从含有109个突变体的文库中成功筛选到12株对抗生素抗性显著提高的阳性突变体.目前,构建数目庞大的突变体文库的技术已日趋成熟(表1列举了不同诱变手段产生的不同数量级大小的突变文库),而最大瓶颈是缺乏与之很好匹配的高通量筛选方法.因此,本文作者将对近些年已经应用或未来有应用潜力的高通量筛选方法进行归类综述,以便为后续研究者们的理性设计和选择提供重要依据和参考. ...
High-throughput screening of antibiotic-resistant bacteria in picodroplets
1
2016
... 菌种进化工程成功的关键是获得多样性的突变文库和开发高通量的筛选方法(图1).突变文库越大,突变体数量越多,则筛选到目标表型菌株的可能性就会越大.而对于庞大的突变体文库,高通量的筛选方法的开发是至关重要的[6],在一项研究中,Liu等[7]利用液滴微流控技术在短短的13h内从含有109个突变体的文库中成功筛选到12株对抗生素抗性显著提高的阳性突变体.目前,构建数目庞大的突变体文库的技术已日趋成熟(表1列举了不同诱变手段产生的不同数量级大小的突变文库),而最大瓶颈是缺乏与之很好匹配的高通量筛选方法.因此,本文作者将对近些年已经应用或未来有应用潜力的高通量筛选方法进行归类综述,以便为后续研究者们的理性设计和选择提供重要依据和参考. ...
A fluorescence-activated cell sorting-based strategy for rapid isolation of high-lipid C hlamydomonas mutants
1
2015
... 菌种进化工程产生的突变文库及其筛选方法
| 菌株 | 进化手段 | 进化目的 | 突变文库大小 | 筛选方法 | 参考文献 |
|---|
| 莱茵衣藻 | 基因重组 | 提高细胞脂质积累 | 6×104 | 基于细胞颜色筛选 | [8] |
| 酵母 | UV诱变 | 提高菌株α-淀粉酶的分泌能力 | 105 | 液滴微流体平台筛选 | [9] |
| 谷氨酸棒杆菌 | ARTP诱变 | 提升L-丝氨酸的生产能力 | 1.2 × 105 | 生物传感器筛选 | [10] |
| 莱茵衣藻 | 甲磺酸乙酯化学诱变 | 增加细胞生长和脂质积累 | 2×105 | 液滴微流体平台筛选 | [11] |
| 大肠杆菌 | ARTP诱变 | 提高菌株L-赖氨酸的生产能力 | 107 | 生物传感器筛选 | [12] |
| 大肠杆菌 | ARTP诱变 | 提高菌株苏氨酸的生产能力 | 2 × 107 | 生物传感器筛选 | [13] |
注: ARTP诱变为常压室温等离子体诱变. ...
High-throughput screening for industrial enzyme production hosts by droplet microfluidics
2
2014
... 菌种进化工程产生的突变文库及其筛选方法
| 菌株 | 进化手段 | 进化目的 | 突变文库大小 | 筛选方法 | 参考文献 |
|---|
| 莱茵衣藻 | 基因重组 | 提高细胞脂质积累 | 6×104 | 基于细胞颜色筛选 | [8] |
| 酵母 | UV诱变 | 提高菌株α-淀粉酶的分泌能力 | 105 | 液滴微流体平台筛选 | [9] |
| 谷氨酸棒杆菌 | ARTP诱变 | 提升L-丝氨酸的生产能力 | 1.2 × 105 | 生物传感器筛选 | [10] |
| 莱茵衣藻 | 甲磺酸乙酯化学诱变 | 增加细胞生长和脂质积累 | 2×105 | 液滴微流体平台筛选 | [11] |
| 大肠杆菌 | ARTP诱变 | 提高菌株L-赖氨酸的生产能力 | 107 | 生物传感器筛选 | [12] |
| 大肠杆菌 | ARTP诱变 | 提高菌株苏氨酸的生产能力 | 2 × 107 | 生物传感器筛选 | [13] |
注: ARTP诱变为常压室温等离子体诱变. ...
... 宏观尺度上代谢物的检测通常使用气质色谱或液质色谱这些大型仪器,虽然能够实现高精度、更广泛的代谢物检测,但是仪器的使用和维护成本非常高,而且样品制备耗时,筛选效率低下.液滴微流体技术的发展为代谢物的检测提供了新的途径,不仅可以实现更快的样品制备,而且液滴微反应器具有对单细胞进行分离区室化的功能,使不同突变的细胞可以在液滴微反应器中进行独立的培养,并且产生的代谢物在各自的微反应器中被隔开而不会相互干扰,因此在目标代谢物高产菌株的筛选中具有广阔的应用前景.如在对工业酶的生产菌株优化研究中,Sjostrom等[9]运用紫外诱变技术对表达α-淀粉酶的酵母菌株进行诱变,并且利用液滴微流体技术平台从含有105个突变体文库中筛选出一株α-淀粉酶表达量提高两倍的稳定高产菌株,该平台的筛选效率比自动微量滴定板筛选系统高出300倍,同时,筛选实验的试剂消耗量减少了1百万倍,大大降低了生产菌株筛选的成本.随后,Huang等[72]同样应用液滴微流体高通量筛选技术,经过多轮筛选,选择了8株α-淀粉酶分泌明显提高的菌株.通过对所有8株优良突变体进行全基因组测序,发现了330处基因突变,对突变的分析揭示了之前未曾发现的与蛋白分泌有关的生理机制,这将有助于研究人员设计更加高效的蛋白分泌细胞工厂,促进合成途径的理性构建.基于小分子代谢物生产和底物消耗对微生物菌株进行表型筛选是许多生物技术关键的瓶颈,Wang等[73]利用液滴微流体平台开发了一套高效筛选方法,用于对酿酒酵母木糖消耗和大肠杆菌乳酸生产的优良表型菌株的筛选.研究人员将荧光探针和单细胞菌株包裹在微反应器中,控制相同的孵育培养条件,通过对荧光信号的检测分析,能够评估微反应器里木糖或者乳酸浓度的变化,进而获得了理想表型的优良菌株. ...
Integration of ARTP mutagenesis with biosensor-mediated high-throughput screening to improve l-serine yield in Corynebacterium glutamicum
1
2018
... 菌种进化工程产生的突变文库及其筛选方法
| 菌株 | 进化手段 | 进化目的 | 突变文库大小 | 筛选方法 | 参考文献 |
|---|
| 莱茵衣藻 | 基因重组 | 提高细胞脂质积累 | 6×104 | 基于细胞颜色筛选 | [8] |
| 酵母 | UV诱变 | 提高菌株α-淀粉酶的分泌能力 | 105 | 液滴微流体平台筛选 | [9] |
| 谷氨酸棒杆菌 | ARTP诱变 | 提升L-丝氨酸的生产能力 | 1.2 × 105 | 生物传感器筛选 | [10] |
| 莱茵衣藻 | 甲磺酸乙酯化学诱变 | 增加细胞生长和脂质积累 | 2×105 | 液滴微流体平台筛选 | [11] |
| 大肠杆菌 | ARTP诱变 | 提高菌株L-赖氨酸的生产能力 | 107 | 生物传感器筛选 | [12] |
| 大肠杆菌 | ARTP诱变 | 提高菌株苏氨酸的生产能力 | 2 × 107 | 生物传感器筛选 | [13] |
注: ARTP诱变为常压室温等离子体诱变. ...
High-throughput droplet microfluidics screening platform for selecting fast-growing and high lipid-producing microalgae from a mutant library
1
2017
... 菌种进化工程产生的突变文库及其筛选方法
| 菌株 | 进化手段 | 进化目的 | 突变文库大小 | 筛选方法 | 参考文献 |
|---|
| 莱茵衣藻 | 基因重组 | 提高细胞脂质积累 | 6×104 | 基于细胞颜色筛选 | [8] |
| 酵母 | UV诱变 | 提高菌株α-淀粉酶的分泌能力 | 105 | 液滴微流体平台筛选 | [9] |
| 谷氨酸棒杆菌 | ARTP诱变 | 提升L-丝氨酸的生产能力 | 1.2 × 105 | 生物传感器筛选 | [10] |
| 莱茵衣藻 | 甲磺酸乙酯化学诱变 | 增加细胞生长和脂质积累 | 2×105 | 液滴微流体平台筛选 | [11] |
| 大肠杆菌 | ARTP诱变 | 提高菌株L-赖氨酸的生产能力 | 107 | 生物传感器筛选 | [12] |
| 大肠杆菌 | ARTP诱变 | 提高菌株苏氨酸的生产能力 | 2 × 107 | 生物传感器筛选 | [13] |
注: ARTP诱变为常压室温等离子体诱变. ...
Evolving the L-lysine high-producing strain of Escherichia coli using a newly developed high-throughput screening method
1
2016
... 菌种进化工程产生的突变文库及其筛选方法
| 菌株 | 进化手段 | 进化目的 | 突变文库大小 | 筛选方法 | 参考文献 |
|---|
| 莱茵衣藻 | 基因重组 | 提高细胞脂质积累 | 6×104 | 基于细胞颜色筛选 | [8] |
| 酵母 | UV诱变 | 提高菌株α-淀粉酶的分泌能力 | 105 | 液滴微流体平台筛选 | [9] |
| 谷氨酸棒杆菌 | ARTP诱变 | 提升L-丝氨酸的生产能力 | 1.2 × 105 | 生物传感器筛选 | [10] |
| 莱茵衣藻 | 甲磺酸乙酯化学诱变 | 增加细胞生长和脂质积累 | 2×105 | 液滴微流体平台筛选 | [11] |
| 大肠杆菌 | ARTP诱变 | 提高菌株L-赖氨酸的生产能力 | 107 | 生物传感器筛选 | [12] |
| 大肠杆菌 | ARTP诱变 | 提高菌株苏氨酸的生产能力 | 2 × 107 | 生物传感器筛选 | [13] |
注: ARTP诱变为常压室温等离子体诱变. ...
Developing a high-throughput screening method for threonine overproduction based on an artificial promoter
1
2015
... 菌种进化工程产生的突变文库及其筛选方法
| 菌株 | 进化手段 | 进化目的 | 突变文库大小 | 筛选方法 | 参考文献 |
|---|
| 莱茵衣藻 | 基因重组 | 提高细胞脂质积累 | 6×104 | 基于细胞颜色筛选 | [8] |
| 酵母 | UV诱变 | 提高菌株α-淀粉酶的分泌能力 | 105 | 液滴微流体平台筛选 | [9] |
| 谷氨酸棒杆菌 | ARTP诱变 | 提升L-丝氨酸的生产能力 | 1.2 × 105 | 生物传感器筛选 | [10] |
| 莱茵衣藻 | 甲磺酸乙酯化学诱变 | 增加细胞生长和脂质积累 | 2×105 | 液滴微流体平台筛选 | [11] |
| 大肠杆菌 | ARTP诱变 | 提高菌株L-赖氨酸的生产能力 | 107 | 生物传感器筛选 | [12] |
| 大肠杆菌 | ARTP诱变 | 提高菌株苏氨酸的生产能力 | 2 × 107 | 生物传感器筛选 | [13] |
注: ARTP诱变为常压室温等离子体诱变. ...
Construction of lycopene-overproducing E. coli strains by combining systematic and combinatorial gene knockout targets
1
2005
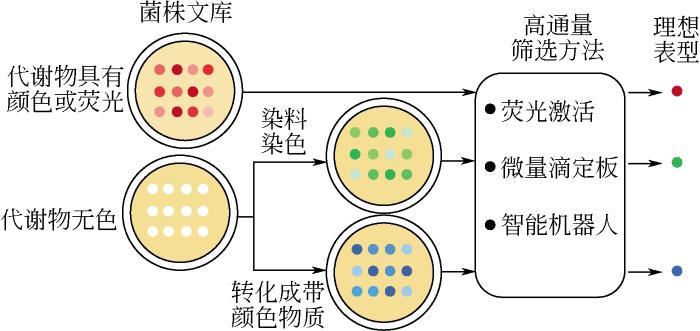
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
Discovery of several novel targets that enhance β-carotene production in Saccharomyces cerevisiae
1
2017
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
Alleviation of metabolic bottleneck by combinatorial engineering enhanced astaxanthin synthesis in Saccharomyces cerevisiae
1
2017
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
Combinatorial and high-throughput screening approaches for strain engineering
1
2015
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
High-throughput metabolic engineering: advances in small-molecule screening and selection
1
2010
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
Looking for the pick of the bunch: high-throughput screening of producing microorganisms with biosensors
1
2014
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
Efficient screening for astaxanthin-overproducing mutants of the yeast Xanthophyllomyces dendrorhous by flow cytometry
1
2008
... 基于颜色或荧光对细胞进行筛选是一种非常直观的高通量筛选方法(图2),在菌种进化工程中广泛而有效地应用于筛选具有颜色或者荧光代谢物菌株.对于生产有色产物(如番茄红素[14]、β-胡萝卜素[15]和虾青素[16])的菌株,颜色的类别和深浅程度可以初步判断反应代谢物的种类和产量高低,通过现代机器人技术辅助作用,筛选效率可以达到每次106个突变体[17].另一种是采用微量滴定板筛选的方法,将菌株代谢物产量与光度测定相关联,以确定突变菌株的效果,每次能够筛选105个突变体[18].近些年荧光激活细胞分选技术(fluorescence-activated cell sorting,FACS)的发展大幅提高了筛选效率,对于细胞内带有荧光的代谢物或者可以被荧光染色的物质,可以设置特定波长的激发光激活细胞内荧光信号,在单细胞水平上根据荧光强度对细胞进行分选,每次筛选通量能够达到109个突变体[19].例如,Ukibe等[20]利用荧光激活细胞分选技术,仅用1h就从超过106个突变体的文库中成功分离出虾青素生产改良的菌株细胞,筛选效率比传统微量滴定板高出100倍. ...
Quantified high-throughput screening of Escherichia coli producing poly(3-hydroxybutyrate) based on FACS
1
2013
... 另外,如果代谢物本身是无色或者缺乏荧光,还可以采用如下改进方案进行筛选.①使用可与代谢物反应产生颜色或荧光的添加试剂.例如,Lee等[21]使氟硼二吡咯与聚-3-羟基丁酸酯反应产生荧光,并且结合高速流式细胞分析仪,实现了对大肠杆菌内聚-3-羟基丁酸酯更高灵敏度的检测.②引入外源酶将细胞代谢物转化成有颜色的化学物质.例如,Deloache等[22]在酿酒酵母体内引入来自植物的二羟基苯丙氨酸双加氧酶(dihydroxyphenylalanine dioxygenase,DOD),它能够将L-多巴(L-3,4-dihydroxyphenylalanine,L-DOPA) 转化为带有黄色荧光的甜菜黄素,并且发现黄色荧光强度与细胞内L-多巴的浓度具有较强的相关性,从而可以通过肉眼直接进行分辨筛选L-多巴高产菌株.运用这种筛选策略,研究人员快速地从突变文库中筛选到一株L-多巴产量比野生菌株提高2.8倍的阳性突变体.同时该策略的建立也为生产无颜色代谢物细胞工厂的筛选提供了高效可行的筛选方法. ...
An enzyme-coupled biosensor enables (S)-reticuline production in yeast from glucose
1
2015
... 另外,如果代谢物本身是无色或者缺乏荧光,还可以采用如下改进方案进行筛选.①使用可与代谢物反应产生颜色或荧光的添加试剂.例如,Lee等[21]使氟硼二吡咯与聚-3-羟基丁酸酯反应产生荧光,并且结合高速流式细胞分析仪,实现了对大肠杆菌内聚-3-羟基丁酸酯更高灵敏度的检测.②引入外源酶将细胞代谢物转化成有颜色的化学物质.例如,Deloache等[22]在酿酒酵母体内引入来自植物的二羟基苯丙氨酸双加氧酶(dihydroxyphenylalanine dioxygenase,DOD),它能够将L-多巴(L-3,4-dihydroxyphenylalanine,L-DOPA) 转化为带有黄色荧光的甜菜黄素,并且发现黄色荧光强度与细胞内L-多巴的浓度具有较强的相关性,从而可以通过肉眼直接进行分辨筛选L-多巴高产菌株.运用这种筛选策略,研究人员快速地从突变文库中筛选到一株L-多巴产量比野生菌株提高2.8倍的阳性突变体.同时该策略的建立也为生产无颜色代谢物细胞工厂的筛选提供了高效可行的筛选方法. ...
Identification of novel benzoylformate decarboxylases by growth selection
1
2006
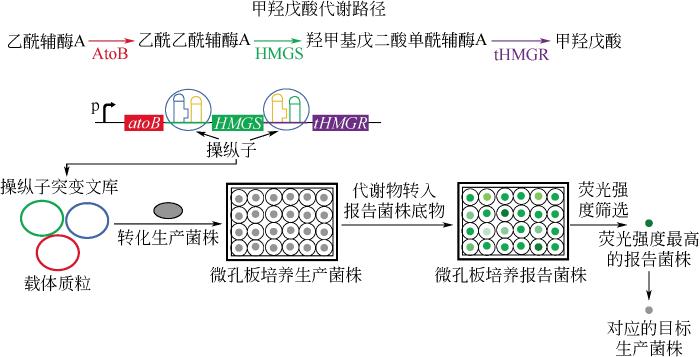
... 基于细胞生长的筛选技术是使用营养缺陷型菌株作为报告系统,用于代谢物高产菌株或者特定酶的筛选.营养型缺陷菌株丧失了合成某一种自身生长必须物质的能力,它们在普通培养基里不能生长,必须补充特定的营养物质,因此可以用来对合成这种必须成分的酶或者代谢路径进行高通量筛选,如苯甲酰甲酸脱羧酶[23]、甲羟戊酸路径[24]、脂肪酶A[25]、糖转运蛋白[26].Pfleger等[24]利用突变调控元件的方法来平衡甲羟戊酸路径的3个酶的相对表达量,以提升甲羟戊酸产量,随后研究人员构建了营养缺陷型报告菌株对突变文库进行筛选(图3).报告菌株依赖甲羟戊酸生长,并表达绿色荧光蛋白基因.生产甲羟戊酸的菌株经过24h培养后离心除去细胞,并将其上清液用作报告菌株的底物.当甲羟戊酸被合成时,细胞方能在培养基上生长,且甲羟戊酸的产量可通过细胞的荧光强度来监测,最终从突变体文库中成功筛选到了甲羟戊酸产量增加7倍的高产突变体.自然界中的酶通常缺乏对底物的特异选择性,阻碍了其在工业上的应用,Boersma等[25]采用进化的方法对枯草芽孢杆菌中的脂肪酶 A进行改造,并选择天冬氨酸营养缺陷型大肠杆菌作为宿主用于脂肪酶 A进化文库的筛选.S-天冬氨酸酯作为选择性底物加入基本培养基,当底物能被脂肪酶 A 水解生成天冬氨酸时,大肠杆菌方可在培养基中生长.为了进一步抑制低选择性的突变体,进化过程中添加可与对映体R-天冬氨酸酯共价结合的抑制剂,并逐渐增加抑制剂浓度增加营养压力,经过3轮迭代筛选,最后成功获得一种对S-天冬氨酸酯的立体选择性显著提高的突变体. ...
Combinatorial engineering of intergenic regions in operons tunes expression of multiple genes
2
2006
... 基于细胞生长的筛选技术是使用营养缺陷型菌株作为报告系统,用于代谢物高产菌株或者特定酶的筛选.营养型缺陷菌株丧失了合成某一种自身生长必须物质的能力,它们在普通培养基里不能生长,必须补充特定的营养物质,因此可以用来对合成这种必须成分的酶或者代谢路径进行高通量筛选,如苯甲酰甲酸脱羧酶[23]、甲羟戊酸路径[24]、脂肪酶A[25]、糖转运蛋白[26].Pfleger等[24]利用突变调控元件的方法来平衡甲羟戊酸路径的3个酶的相对表达量,以提升甲羟戊酸产量,随后研究人员构建了营养缺陷型报告菌株对突变文库进行筛选(图3).报告菌株依赖甲羟戊酸生长,并表达绿色荧光蛋白基因.生产甲羟戊酸的菌株经过24h培养后离心除去细胞,并将其上清液用作报告菌株的底物.当甲羟戊酸被合成时,细胞方能在培养基上生长,且甲羟戊酸的产量可通过细胞的荧光强度来监测,最终从突变体文库中成功筛选到了甲羟戊酸产量增加7倍的高产突变体.自然界中的酶通常缺乏对底物的特异选择性,阻碍了其在工业上的应用,Boersma等[25]采用进化的方法对枯草芽孢杆菌中的脂肪酶 A进行改造,并选择天冬氨酸营养缺陷型大肠杆菌作为宿主用于脂肪酶 A进化文库的筛选.S-天冬氨酸酯作为选择性底物加入基本培养基,当底物能被脂肪酶 A 水解生成天冬氨酸时,大肠杆菌方可在培养基中生长.为了进一步抑制低选择性的突变体,进化过程中添加可与对映体R-天冬氨酸酯共价结合的抑制剂,并逐渐增加抑制剂浓度增加营养压力,经过3轮迭代筛选,最后成功获得一种对S-天冬氨酸酯的立体选择性显著提高的突变体. ...
... [24]利用突变调控元件的方法来平衡甲羟戊酸路径的3个酶的相对表达量,以提升甲羟戊酸产量,随后研究人员构建了营养缺陷型报告菌株对突变文库进行筛选(图3).报告菌株依赖甲羟戊酸生长,并表达绿色荧光蛋白基因.生产甲羟戊酸的菌株经过24h培养后离心除去细胞,并将其上清液用作报告菌株的底物.当甲羟戊酸被合成时,细胞方能在培养基上生长,且甲羟戊酸的产量可通过细胞的荧光强度来监测,最终从突变体文库中成功筛选到了甲羟戊酸产量增加7倍的高产突变体.自然界中的酶通常缺乏对底物的特异选择性,阻碍了其在工业上的应用,Boersma等[25]采用进化的方法对枯草芽孢杆菌中的脂肪酶 A进行改造,并选择天冬氨酸营养缺陷型大肠杆菌作为宿主用于脂肪酶 A进化文库的筛选.S-天冬氨酸酯作为选择性底物加入基本培养基,当底物能被脂肪酶 A 水解生成天冬氨酸时,大肠杆菌方可在培养基中生长.为了进一步抑制低选择性的突变体,进化过程中添加可与对映体R-天冬氨酸酯共价结合的抑制剂,并逐渐增加抑制剂浓度增加营养压力,经过3轮迭代筛选,最后成功获得一种对S-天冬氨酸酯的立体选择性显著提高的突变体. ...
A novel genetic selection system for improved enantioselectivity of Bacillus subtilis lipase A
2
2008
... 基于细胞生长的筛选技术是使用营养缺陷型菌株作为报告系统,用于代谢物高产菌株或者特定酶的筛选.营养型缺陷菌株丧失了合成某一种自身生长必须物质的能力,它们在普通培养基里不能生长,必须补充特定的营养物质,因此可以用来对合成这种必须成分的酶或者代谢路径进行高通量筛选,如苯甲酰甲酸脱羧酶[23]、甲羟戊酸路径[24]、脂肪酶A[25]、糖转运蛋白[26].Pfleger等[24]利用突变调控元件的方法来平衡甲羟戊酸路径的3个酶的相对表达量,以提升甲羟戊酸产量,随后研究人员构建了营养缺陷型报告菌株对突变文库进行筛选(图3).报告菌株依赖甲羟戊酸生长,并表达绿色荧光蛋白基因.生产甲羟戊酸的菌株经过24h培养后离心除去细胞,并将其上清液用作报告菌株的底物.当甲羟戊酸被合成时,细胞方能在培养基上生长,且甲羟戊酸的产量可通过细胞的荧光强度来监测,最终从突变体文库中成功筛选到了甲羟戊酸产量增加7倍的高产突变体.自然界中的酶通常缺乏对底物的特异选择性,阻碍了其在工业上的应用,Boersma等[25]采用进化的方法对枯草芽孢杆菌中的脂肪酶 A进行改造,并选择天冬氨酸营养缺陷型大肠杆菌作为宿主用于脂肪酶 A进化文库的筛选.S-天冬氨酸酯作为选择性底物加入基本培养基,当底物能被脂肪酶 A 水解生成天冬氨酸时,大肠杆菌方可在培养基中生长.为了进一步抑制低选择性的突变体,进化过程中添加可与对映体R-天冬氨酸酯共价结合的抑制剂,并逐渐增加抑制剂浓度增加营养压力,经过3轮迭代筛选,最后成功获得一种对S-天冬氨酸酯的立体选择性显著提高的突变体. ...
... [25]采用进化的方法对枯草芽孢杆菌中的脂肪酶 A进行改造,并选择天冬氨酸营养缺陷型大肠杆菌作为宿主用于脂肪酶 A进化文库的筛选.S-天冬氨酸酯作为选择性底物加入基本培养基,当底物能被脂肪酶 A 水解生成天冬氨酸时,大肠杆菌方可在培养基中生长.为了进一步抑制低选择性的突变体,进化过程中添加可与对映体R-天冬氨酸酯共价结合的抑制剂,并逐渐增加抑制剂浓度增加营养压力,经过3轮迭代筛选,最后成功获得一种对S-天冬氨酸酯的立体选择性显著提高的突变体. ...
A growth-based screening system for hexose transporters in yeast
1
2018
... 基于细胞生长的筛选技术是使用营养缺陷型菌株作为报告系统,用于代谢物高产菌株或者特定酶的筛选.营养型缺陷菌株丧失了合成某一种自身生长必须物质的能力,它们在普通培养基里不能生长,必须补充特定的营养物质,因此可以用来对合成这种必须成分的酶或者代谢路径进行高通量筛选,如苯甲酰甲酸脱羧酶[23]、甲羟戊酸路径[24]、脂肪酶A[25]、糖转运蛋白[26].Pfleger等[24]利用突变调控元件的方法来平衡甲羟戊酸路径的3个酶的相对表达量,以提升甲羟戊酸产量,随后研究人员构建了营养缺陷型报告菌株对突变文库进行筛选(图3).报告菌株依赖甲羟戊酸生长,并表达绿色荧光蛋白基因.生产甲羟戊酸的菌株经过24h培养后离心除去细胞,并将其上清液用作报告菌株的底物.当甲羟戊酸被合成时,细胞方能在培养基上生长,且甲羟戊酸的产量可通过细胞的荧光强度来监测,最终从突变体文库中成功筛选到了甲羟戊酸产量增加7倍的高产突变体.自然界中的酶通常缺乏对底物的特异选择性,阻碍了其在工业上的应用,Boersma等[25]采用进化的方法对枯草芽孢杆菌中的脂肪酶 A进行改造,并选择天冬氨酸营养缺陷型大肠杆菌作为宿主用于脂肪酶 A进化文库的筛选.S-天冬氨酸酯作为选择性底物加入基本培养基,当底物能被脂肪酶 A 水解生成天冬氨酸时,大肠杆菌方可在培养基中生长.为了进一步抑制低选择性的突变体,进化过程中添加可与对映体R-天冬氨酸酯共价结合的抑制剂,并逐渐增加抑制剂浓度增加营养压力,经过3轮迭代筛选,最后成功获得一种对S-天冬氨酸酯的立体选择性显著提高的突变体. ...
Applications of genetically-encoded biosensors for the construction and control of biosynthetic pathways
1
2012
... 在微生物生产的化学品中,只有少数可以通过颜色或者荧光的筛选方法直接进行筛选.许多化学物质本身不具有颜色或荧光,甚至难以转化为易于染色或者有颜色的物质.在微生物体内广泛存在一类蛋白质或RNA,它们能够识别并响应细胞内特定的代谢物,并转化为特定的信号输出[27](如荧光、细胞的生长、代谢通路的开闭),通过信号强度检测细胞代谢物浓度.基于这一原理,研究人员开发了一系列生物传感器的筛选方法,用于菌种进化工程突变文库的高通量筛选. ...
Transcription factors: an overview
1
1997
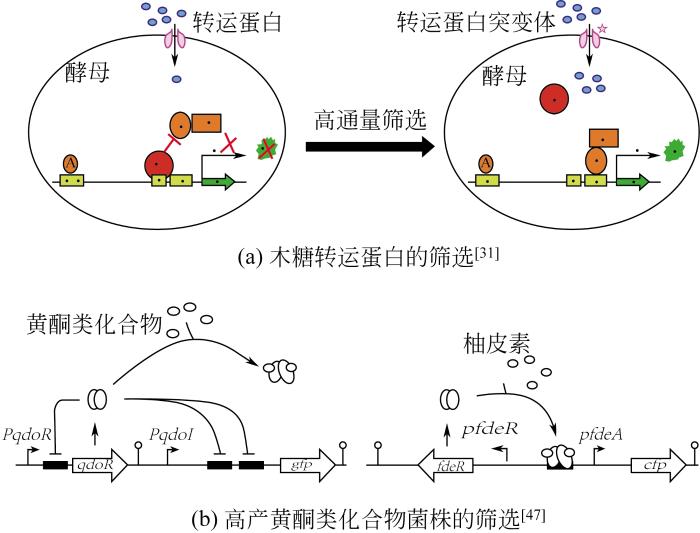
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
A high-throughput approach to identify genomic variants of bacterial metabolite producers at the single-cell level
1
2012
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
Biosensor-driven adaptive laboratory evolution of L-valine production in Corynebacterium glutamicum
2
2015
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Design and engineering of intracellular-metabolite-sensing/regulation gene circuits in Saccharomyces cerevisiae
2
2016
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Programming adaptive control to evolve increased metabolite production
2
2013
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Transcription factor-based screens and synthetic selections for microbial small-molecule biosynthesis
2
2012
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Evolution-guided optimization of biosynthetic pathways
2
2014
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
So x R as a single-cell biosensor for NADPH-consuming enzymes in Escherichia coli
2
2013
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
36(suppl1): D93-D96
1
2007
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
36(suppl1): D120-D124
1
2008
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
34(s1): D108-D110
1
2006
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
Transcription regulatory regions database (TRRD): its status in 2002
1
2002
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
RegTransBase——a database of regulatory sequences and interactions based on literature: a resource for investigating transcriptional regulation in prokaryotes
1
2013
... 转录因子能够结合在基因的启动子或者增强子区域,独自或与其他蛋白组成复合体,进而促进或者阻断RNA聚合酶参与转录过程,来调控基因表达[28].细胞内一些小分子代谢物可以通过配体结合效应、磷酸化作用,激活或者失活转录因子,控制相关报告基因的表达,从而将代谢物浓度与荧光强度、细胞生长等检测信号联系起来,达到高通量筛选目的.响应目标代谢物的转录因子目前主要分为两类:天然转录因子和人工转录因子.在自然界微生物体内,响应小分子的天然转录因子广泛存在.其中在大肠杆菌中发现了230多种转录因子,它们能够感知各种各样的代谢物,包括氨基酸、糖类、糖磷酸和脂质等[29].目前,研究者可以利用天然转录因子开发响应不同代谢物的高通量筛选生物传感器.Regina等[30]利用谷氨酸棒杆菌天然转录调节因子Lrp在谷氨酸棒杆菌体内设计构建了一种响应L-缬氨酸的生物传感器,L-缬氨酸的浓度转化为不同强度的荧光信号输出,在进化压力条件下,使用荧光激活细胞分选技术从进化文库中筛选出具有最高荧光强度的突变菌株,并进行连续迭代培养和分离,最终成功获得了优良的突变体,表现出更快的生长速率并将L-缬氨酸的产量提升了25%,副产物减少了3~4倍.这一研究成果显示出天然转录因子在生物传感器中的巨大应用潜力,同时为菌种进化工程改善微生物菌株的生长和生产效率提供了更加高效的筛选方法.目前,研究者利用天然转录因子,已开发了不同的生物传感器,可以响应木糖[31]、酪氨酸[32]、1-丁醇[33]、葡萄糖二酸和柚皮素[34]以及乙醇脱氢酶[35].随着生物信息学技术的快速发展,研究人员开发了很多转录因子调控的数据库,如DBTBS[36]、RegulonDB[37]、TRANSFAC[38]、TRRD[39]、RegTransBase[40]等,这些数据库总结了各种微生物体内的的转录因子及其识别调控区域和响应的小分子,为生物传感器的开发提供了强有力的信息支撑. ...
Analysis of amino acid substitutions in AraC variants that respond to triacetic acid lactone
2
2016
... 到目前为止,大多数基于转录因子的生物传感器主要依赖于天然转录因子,然而天然转录因子对代谢物的响应具有高度的特异性,在实际应用中,微生物细胞工厂生产的目标代谢物往往不存在天然响应的转录因子,为了实现更广泛的利用,可以通过工程化的手段改变转录因子响应代谢物的特异性.例如,研究人员运用各种工程化的方法对大肠杆菌中的响应L-阿拉伯糖的天然转录因子进行改造,使其特异性发生改变,响应代谢物范围充分扩展,已经能够特异性响应三乙酸内酯[41,42]、甲羟戊酸[43]、D-阿拉伯糖[44]和四氢嘧啶[45].基于转录因子的高通量筛选及应用见表2及图4. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Screening for enhanced triacetic acid lactone production by recombinant Escherichia coli expressing a designed triacetic acid lactone reporter
2
2013
... 到目前为止,大多数基于转录因子的生物传感器主要依赖于天然转录因子,然而天然转录因子对代谢物的响应具有高度的特异性,在实际应用中,微生物细胞工厂生产的目标代谢物往往不存在天然响应的转录因子,为了实现更广泛的利用,可以通过工程化的手段改变转录因子响应代谢物的特异性.例如,研究人员运用各种工程化的方法对大肠杆菌中的响应L-阿拉伯糖的天然转录因子进行改造,使其特异性发生改变,响应代谢物范围充分扩展,已经能够特异性响应三乙酸内酯[41,42]、甲羟戊酸[43]、D-阿拉伯糖[44]和四氢嘧啶[45].基于转录因子的高通量筛选及应用见表2及图4. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Design and application of a mevalonate-responsive regulatory protein
2
2011
... 到目前为止,大多数基于转录因子的生物传感器主要依赖于天然转录因子,然而天然转录因子对代谢物的响应具有高度的特异性,在实际应用中,微生物细胞工厂生产的目标代谢物往往不存在天然响应的转录因子,为了实现更广泛的利用,可以通过工程化的手段改变转录因子响应代谢物的特异性.例如,研究人员运用各种工程化的方法对大肠杆菌中的响应L-阿拉伯糖的天然转录因子进行改造,使其特异性发生改变,响应代谢物范围充分扩展,已经能够特异性响应三乙酸内酯[41,42]、甲羟戊酸[43]、D-阿拉伯糖[44]和四氢嘧啶[45].基于转录因子的高通量筛选及应用见表2及图4. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
AraC regulatory protein mutants with altered effector specificity
2
2008
... 到目前为止,大多数基于转录因子的生物传感器主要依赖于天然转录因子,然而天然转录因子对代谢物的响应具有高度的特异性,在实际应用中,微生物细胞工厂生产的目标代谢物往往不存在天然响应的转录因子,为了实现更广泛的利用,可以通过工程化的手段改变转录因子响应代谢物的特异性.例如,研究人员运用各种工程化的方法对大肠杆菌中的响应L-阿拉伯糖的天然转录因子进行改造,使其特异性发生改变,响应代谢物范围充分扩展,已经能够特异性响应三乙酸内酯[41,42]、甲羟戊酸[43]、D-阿拉伯糖[44]和四氢嘧啶[45].基于转录因子的高通量筛选及应用见表2及图4. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Design of an ectoine-responsive AraC mutant and its application in metabolic engineering of ectoine biosynthesis
2
2015
... 到目前为止,大多数基于转录因子的生物传感器主要依赖于天然转录因子,然而天然转录因子对代谢物的响应具有高度的特异性,在实际应用中,微生物细胞工厂生产的目标代谢物往往不存在天然响应的转录因子,为了实现更广泛的利用,可以通过工程化的手段改变转录因子响应代谢物的特异性.例如,研究人员运用各种工程化的方法对大肠杆菌中的响应L-阿拉伯糖的天然转录因子进行改造,使其特异性发生改变,响应代谢物范围充分扩展,已经能够特异性响应三乙酸内酯[41,42]、甲羟戊酸[43]、D-阿拉伯糖[44]和四氢嘧啶[45].基于转录因子的高通量筛选及应用见表2及图4. ...
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Development of a synthetic malonyl-CoA sensor in Saccharomyces cerevisiae for intracellular metabolite monitoring and genetic screening
2
2015
... 基于转录因子的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 谷氨酸棒杆菌 | L-缬氨酸 | 目标突变体L-缬氨酸的产量提升了25%,副产物减少了3~4倍 | [30] |
| 酿酒酵母 | 木糖 | 开发了一套木糖转运蛋白高通量筛选方法,并获得了一种优良的突变体,其木糖转运能力提高了6.5倍 | [31] |
| 大肠杆菌 | 酪氨酸 | 目标突变体酪氨酸产量提升5倍 | [32] |
| 大肠杆菌 | 1-丁醇 | 目标突变体1-丁醇产量提升35% | [33] |
| 大肠杆菌 | 葡萄糖二酸和柚皮素 | 目标突变体葡萄糖二酸产量提升22倍,柚皮素产量提升36倍 | [34] |
| 大肠杆菌 | 乙醇脱氢酶 | 乙醇脱氢酶对底物的特异性地得到提高 | [35] |
| 大肠杆菌 | 三乙酸内酯 | 成功突变天然转录因子 AraC特异性响应细胞内三乙酸内酯 | [41,42] |
| 大肠杆菌 | 甲羟戊酸 | 成功突变天然转录因子 AraC特异性响应细胞内甲羟戊酸 | [43] |
| 大肠杆菌 | D-阿拉伯糖 | 成功突变天然转录因子 AraC特异性响应细胞内D-阿拉伯糖信号 | [44] |
| 大肠杆菌 | 四氢嘧啶 | 成功突变天然转录因子 AraC特异性响应细胞内四氢嘧啶,并成功筛选到高产四氢嘧啶的目标突变体 | [45] |
| 酿酒酵母 | 丙二酰辅酶A | 目标突变体丙二酰辅酶A表达提高,下游产物3-羟基丙酸的产量提升了120% | [46] |
真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
... 真核生物调控网络要比原核生物复杂很多,目前对其转录调控网络的认识有限,而且细胞内缺乏内源的多功能转录因子和对特定目标代谢物响应的启动子,导致真核细胞内生物传感器的应用受到极大限制.Li等[46]在酿酒酵母体内引入来自细菌的转录因子FapR及其相应的操纵子FapO,构建了响应细胞内丙二酰辅酶A的生物传感器,该传感器将细胞内丙二酰辅酶A水平转换为不同强度的荧光信号,从而在酿酒酵母中实现更简单、更高效的监测和筛选.研究人员利用这种传感装置成功从全基因组cDNA过表达文库中筛选到了高表达丙二酰辅酶A的突变体,并且发现了两个与产量提升有关的靶标基因.在此重组菌株的基础上,使得下游产物3-羟基丙酸的产量提升了120%,这种生物传感器为真核生物进化文库的筛选提供了高效的筛选方法,可以进一步改善酵母中丙二酰辅酶A衍生化学物质的合成. ...
Novel biosensors based on flavonoid-responsive transcriptional regulators introduced into Escherichia coli
2014
Maestro of regulation: riboswitches orchestrate gene expression at the levels of translation, transcription and mRNA decay
1
2018
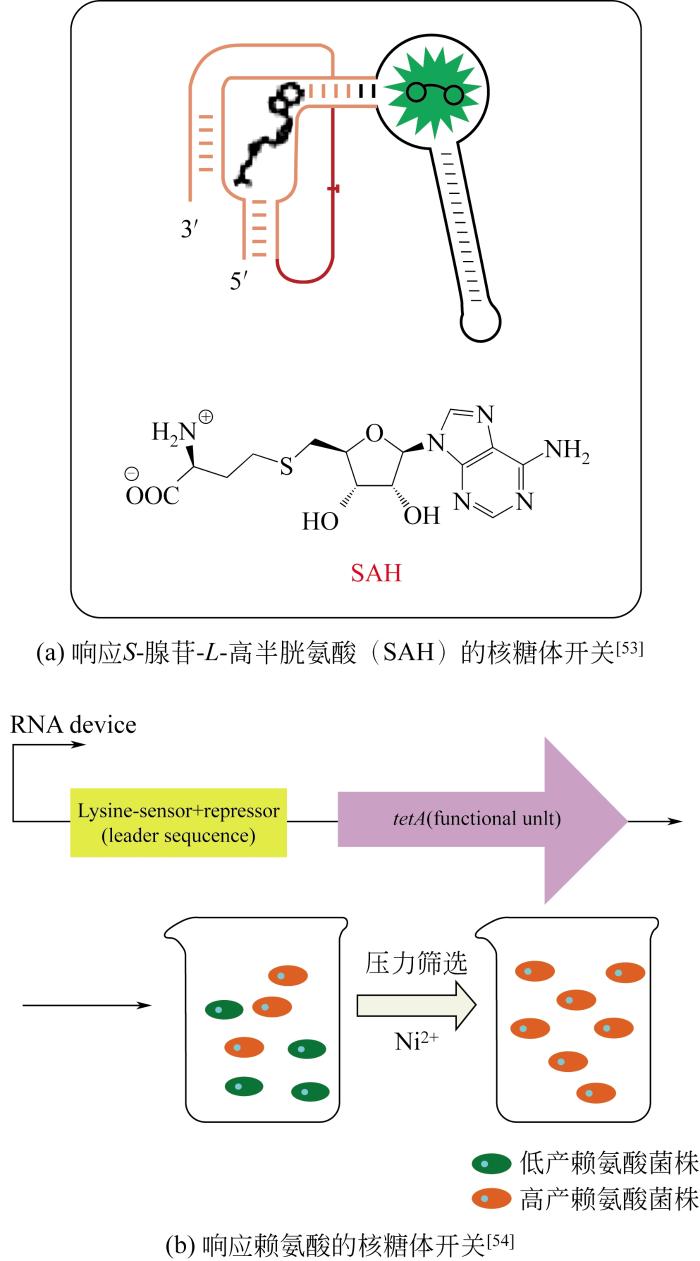
... 核糖开关是基于RNA的基因调控元件,由适配子和基因调控结构域两部分组成.当小分子配体和适配子结合,引起RNA构象发生改变,从而调节基因调控结构域的活性,在转录、翻译和mRNA水平上开启或者关闭蛋白质的合成[48].利用这种调控机制,构建易于检测输出信号的基因模块,同样能够设计响应目标代谢产物的高通量筛选生物传感器(表3,图5). ...
Programmed evolution for optimization of orthogonal metabolic output in bacteria
2
2015
... 基于核糖体开关的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 大肠杆菌 | 茶碱 | 鉴定出了增加茶碱生产的最佳生物元件组合 | [49] |
| 大肠杆菌 | 氰尿二酰胺 | 改造设计正交核糖体开关,不再响应细胞内天然配体分子,而特异性响应非天然小分子氰尿二酰胺 | [50] |
| 酿酒酵母 | 6-磷酸葡糖胺 | 能够有效分离高产n-乙酰氨基葡糖的突变体 | [51] |
| 苜蓿根瘤菌 | VB12 | 获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9% | [52] |
在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
... 在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
Reengineering orthogonally selective riboswitches
2
2010
... 基于核糖体开关的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 大肠杆菌 | 茶碱 | 鉴定出了增加茶碱生产的最佳生物元件组合 | [49] |
| 大肠杆菌 | 氰尿二酰胺 | 改造设计正交核糖体开关,不再响应细胞内天然配体分子,而特异性响应非天然小分子氰尿二酰胺 | [50] |
| 酿酒酵母 | 6-磷酸葡糖胺 | 能够有效分离高产n-乙酰氨基葡糖的突变体 | [51] |
| 苜蓿根瘤菌 | VB12 | 获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9% | [52] |
在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
... 在翻译水平上,核糖体开关感应代谢物并调节mRNA的二级结构,使RBS位点暴露或者隔离,进而促进或者抑制相关报告基因的表达.基于这种机制,合成的核糖开关已被设计用于感知一系列代谢物,包括氰尿二酰胺[50]、6-磷酸葡糖胺[51]、茶碱[57].在一项研究中,Cai等[52]引入来自鼠伤寒沙门氏菌的调控装置第一次在苜蓿根瘤菌体内构建了核糖体开关,用来筛选高产VB12的优良突变菌株(图6).当VB12浓度较高时,核糖体开关RBS序列被隔离,LacI蛋白表达被抑制,绿色荧光蛋白表达增强.当VB12浓度较低时,核糖体开关RBS序列暴露,LacI蛋白正常表达,与LacO操纵子结合抑制绿色荧光蛋白表达.通过建立核糖体开关高通量筛选系统,研究人员成功获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9%. ...
A synthetic suicide riboswitch for the high-throughput screening of metabolite production in Saccharomyces cerevisiae
2
2015
... 基于核糖体开关的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 大肠杆菌 | 茶碱 | 鉴定出了增加茶碱生产的最佳生物元件组合 | [49] |
| 大肠杆菌 | 氰尿二酰胺 | 改造设计正交核糖体开关,不再响应细胞内天然配体分子,而特异性响应非天然小分子氰尿二酰胺 | [50] |
| 酿酒酵母 | 6-磷酸葡糖胺 | 能够有效分离高产n-乙酰氨基葡糖的突变体 | [51] |
| 苜蓿根瘤菌 | VB12 | 获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9% | [52] |
在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
... 在翻译水平上,核糖体开关感应代谢物并调节mRNA的二级结构,使RBS位点暴露或者隔离,进而促进或者抑制相关报告基因的表达.基于这种机制,合成的核糖开关已被设计用于感知一系列代谢物,包括氰尿二酰胺[50]、6-磷酸葡糖胺[51]、茶碱[57].在一项研究中,Cai等[52]引入来自鼠伤寒沙门氏菌的调控装置第一次在苜蓿根瘤菌体内构建了核糖体开关,用来筛选高产VB12的优良突变菌株(图6).当VB12浓度较高时,核糖体开关RBS序列被隔离,LacI蛋白表达被抑制,绿色荧光蛋白表达增强.当VB12浓度较低时,核糖体开关RBS序列暴露,LacI蛋白正常表达,与LacO操纵子结合抑制绿色荧光蛋白表达.通过建立核糖体开关高通量筛选系统,研究人员成功获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9%. ...
Engineering a vitamin B12 high-throughput screening system by riboswitch sensor in Sinorhizobium meliloti
3
2018
... 基于核糖体开关的高通量筛选应用
| 宿主细胞 | 响应物质 | 生物传感器筛选结果或作用 | 参考文献 |
|---|
| 大肠杆菌 | 茶碱 | 鉴定出了增加茶碱生产的最佳生物元件组合 | [49] |
| 大肠杆菌 | 氰尿二酰胺 | 改造设计正交核糖体开关,不再响应细胞内天然配体分子,而特异性响应非天然小分子氰尿二酰胺 | [50] |
| 酿酒酵母 | 6-磷酸葡糖胺 | 能够有效分离高产n-乙酰氨基葡糖的突变体 | [51] |
| 苜蓿根瘤菌 | VB12 | 获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9% | [52] |
在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
... 在翻译水平上,核糖体开关感应代谢物并调节mRNA的二级结构,使RBS位点暴露或者隔离,进而促进或者抑制相关报告基因的表达.基于这种机制,合成的核糖开关已被设计用于感知一系列代谢物,包括氰尿二酰胺[50]、6-磷酸葡糖胺[51]、茶碱[57].在一项研究中,Cai等[52]引入来自鼠伤寒沙门氏菌的调控装置第一次在苜蓿根瘤菌体内构建了核糖体开关,用来筛选高产VB12的优良突变菌株(图6).当VB12浓度较高时,核糖体开关RBS序列被隔离,LacI蛋白表达被抑制,绿色荧光蛋白表达增强.当VB12浓度较低时,核糖体开关RBS序列暴露,LacI蛋白正常表达,与LacO操纵子结合抑制绿色荧光蛋白表达.通过建立核糖体开关高通量筛选系统,研究人员成功获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9%. ...
... [
52]
![]()
目前,基于不同的核糖体开关机制开发的高通量生物传感器数量正在不断增长,但是在代谢工程中应用依然受到限制,主要是缺乏响应特定代谢产物的适配子以及对适配子和驱动结构域的联系机制缺乏了解.近年来,合成生物学的发展为理性设计核糖体开关生物传感器提供了计算机辅助设计工具[58],并且能够对响应调控反应作出预测,未来将有可能极大地促进核糖开关在高通量筛选领域的应用. ...
In vitro and in vivo enzyme activity screening via RNA-based fluorescent biosensors for S-adenosyl-L-homocysteine (SAH)
2016
Evolution of a chimeric aspartate kinase for L-lysine production using a synthetic RNA device
2015
The structure of a tetrahydrofolate-sensing riboswitch reveals two ligand binding sites in a single aptamer
1
2011
... 在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
De novo design of a synthetic riboswitch that regulates transcription termination
1
2012
... 在转录水平上,可以通过工程化的方法改造适配子结合域来促进或者破坏终止子的形成,进而产生转录抑制和激活作用.利用这种机制,已经设计出了响应四氢叶酸[55]和茶碱[49]的生物传感器.同时,也可以将多个核糖体传感器连接在一起,实现更复杂的功能[56].例如,为了增加信号输出动态范围,可以将相同核糖开关串联,结合在单个转录单元中以增加目标代谢物控制的终止子数量,提高生物传感器响应的灵敏度.也可以将响应不同代谢物的核糖体开关组合在一起,设计响应多个环境小分子信号的布尔逻辑门,用于菌种进化工程代谢长路径的筛选. ...
Synthetic riboswitches that induce gene expression in diverse bacterial species
1
2010
... 在翻译水平上,核糖体开关感应代谢物并调节mRNA的二级结构,使RBS位点暴露或者隔离,进而促进或者抑制相关报告基因的表达.基于这种机制,合成的核糖开关已被设计用于感知一系列代谢物,包括氰尿二酰胺[50]、6-磷酸葡糖胺[51]、茶碱[57].在一项研究中,Cai等[52]引入来自鼠伤寒沙门氏菌的调控装置第一次在苜蓿根瘤菌体内构建了核糖体开关,用来筛选高产VB12的优良突变菌株(图6).当VB12浓度较高时,核糖体开关RBS序列被隔离,LacI蛋白表达被抑制,绿色荧光蛋白表达增强.当VB12浓度较低时,核糖体开关RBS序列暴露,LacI蛋白正常表达,与LacO操纵子结合抑制绿色荧光蛋白表达.通过建立核糖体开关高通量筛选系统,研究人员成功获得了一株优良的阳性突变体,VB12产量比野生型提高了21.9%. ...
Automated physics-based design of synthetic riboswitches from diverse RNA aptamers
1
2015
... 目前,基于不同的核糖体开关机制开发的高通量生物传感器数量正在不断增长,但是在代谢工程中应用依然受到限制,主要是缺乏响应特定代谢产物的适配子以及对适配子和驱动结构域的联系机制缺乏了解.近年来,合成生物学的发展为理性设计核糖体开关生物传感器提供了计算机辅助设计工具[58],并且能够对响应调控反应作出预测,未来将有可能极大地促进核糖开关在高通量筛选领域的应用. ...
A genetically encoded F?rster resonance energy transfer sensor for monitoring in vivo trehalose-6-phosphate dynamics
2
2015
... 海藻糖浓度
[59] | 大肠杆菌和 ...
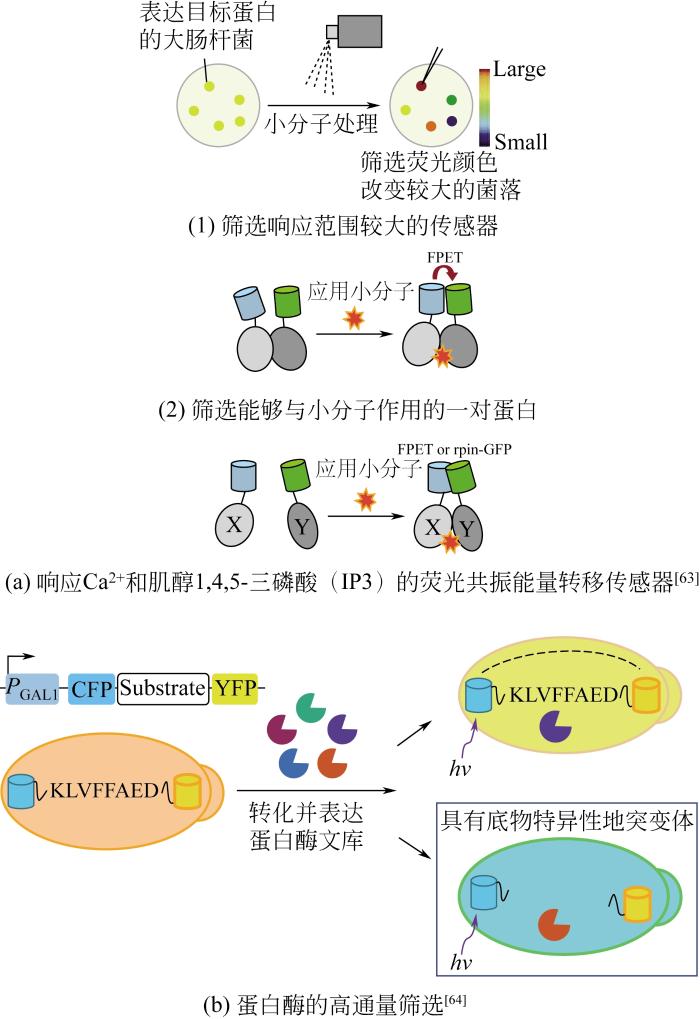
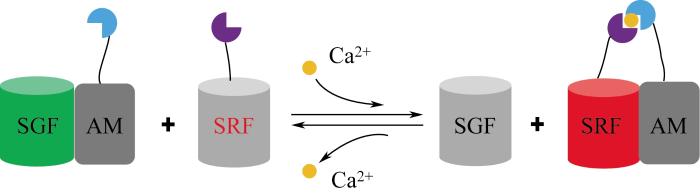
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Genetically-encoded nanosensor for quantitative monitoring of methionine in bacterial and yeast cells
2
2014
| 蛋氨酸 | 细胞内蛋氨酸监测 | [60] |
| 大肠杆菌 | 亮氨酸 | 细胞内亮氨酸监测 | [61] |
| 大肠杆菌 | 抗生素 | 建立了高通量筛选潜在 ...
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Genetically encoded FRET-based nanosensor for in vivo measurement of leucine
2
2013
| 蛋氨酸 | 细胞内蛋氨酸监测 | [60] |
| 大肠杆菌 | 亮氨酸 | 细胞内亮氨酸监测 | [61] |
| 大肠杆菌 | 抗生素 | 建立了高通量筛选潜在 ...
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Engineered EF-Tu and tRNA-based FRET screening assay to find inhibitors of protein synthesis in bacteria
2
2018
| [62] |
| 大肠杆菌 | 肌醇1,4,5-三磷酸 | 建立了高通量筛选小分子 ...
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Development of a convenient and supersensitive high-throughput screening system for genetically encoded fluorescent probes of small molecules using a confocal microscope
1
2017
| [63] |
| 酿酒酵母 | 蛋白酶 | 成功筛选出一种对底物选择性提高30倍的突变体 | [64] |
目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Intracellular FRET-based screen for redesigning the specificity of secreted proteases
1
2016
... 肌醇1,4,5-三磷酸的方法
[63] | | 酿酒酵母 | 蛋白酶 | 成功筛选出一种对底物选择性提高30倍的突变体 | [64] |
目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
In situ monitoring of intracellular glucose and glutamine in CHO cell culture
1
2012
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Novel fluorescence resonance energy transfer optical sensors for vitamin B12 detection using thermally reduced carbon dots
1
2015
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Highly selective and sensitive optosensing of glutathione based on fluorescence resonance energy transfer of upconversion nanoparticles coated with a Rhodamine B derivative
1
2018
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
Ratiometric biosensors based on dimerization-dependent fluorescent protein exchange
2
2015
... 目前基于荧光共振能量转移原理开发的生物传感器能够响应多种重要的代谢物,如 6-磷酸海藻糖[59]、蛋氨酸[60]、 亮氨酸[61]、抗生素[62]、谷氨酰胺[65]、维生素B12[66]和谷胱甘肽[67].在此基础上,为了扩大荧光共振能量转移生物传感器应用的范围和实现更复杂的响应功能,Ding等[68]开发了一种新型荧光共振能量转移生物传感器(图8),用于细胞内Ca2+浓度的监测.该传感器由一对特殊的荧光基团SGF和SRF组成,在单独存在时,两种荧光基团均不发光,只有当与第3种激活单体(activating monomer,AM)竞争性结合时,才会发光.AM和SRF都连接有受体蛋白,当缺乏配体小分子时,AM只能选择性地与SGF结合,发出强烈的绿色荧光.当小分子配体存在时,将AM转移与SRF结合,使能量发生转移,激活红色荧光,从而可以根据红绿荧光比率,检测细胞内目标小分子代谢物的浓度. ...
... [
68]
![]()
尽管基于FRET的双荧光报告系统目前能够响应的小分子越来越多,而且比使用单个荧光基团的生物传感器的精度要高的多,但是信号强度和荧光比率易受到周围环境干扰,比如pH、温度等,目前在高通量筛选领域的应用仍然受到相当多的限制. ...
Microfluidic approaches for systems and synthetic biology
1
2010
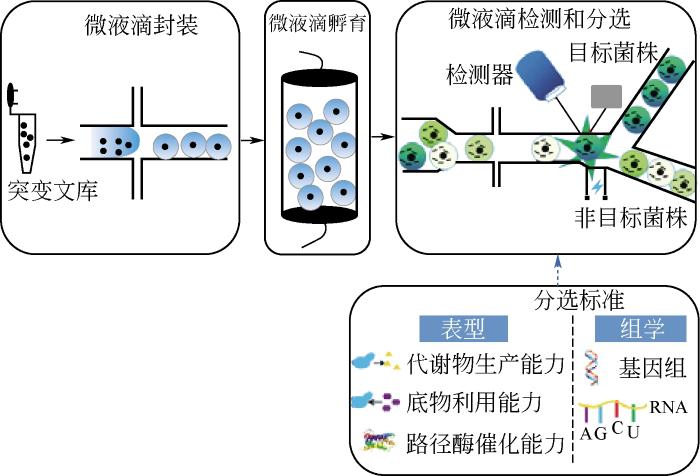
... 液滴微流体平台有单细胞分离良好的隔室,可以作为微反应器进行细胞的恒化培养、蛋白表达分析、代谢物检测和组学分析,为菌种进化工程高通量筛选提供了有利的技术支撑.与传统高通量筛选技术相比,液滴微流体技术表现出更高的通量和精度,大大降低了筛选时间和成本,可以同时分析多达150万个样本[69],并具有快速引入环境扰动的能力,提供动态的微环境条件控制,显著提高了研究人员对微生物体系进行控制和筛选的能力,为菌种进化工程获得理想的微生物菌株提供了有力的工具.因为荧光信号检测具有很高的灵敏度,特别适合作为液滴微流体平台皮升级微反应器的检测信号,所以目前微流控技术主要将代谢物的浓度、路径酶催化能力、组学检测等和荧光信号偶联,通过荧光信号对菌株表型和组学进行筛选(图9). ...
Ultrahigh-throughput screening in drop-based microfluidics for directed evolution
1
2010
... 随着高效率封装单细胞技术的发展,单个细胞能被快速限制在微液滴内,实现细胞内代谢路径酶的高通量筛选.荧光底物被用来检测酶活性,其本身无荧光或者只有很低的荧光信号,但是其酶促反应产物则具有很强的荧光信号,根据反应前后荧光强度的变化,可以对酶活性进行分析和筛选.Agresti等[70]运用进化的方法对辣根过氧化物酶进行突变并转入酵母宿主体内,随后将荧光底物和酵母菌株一起封装在微液滴中,利用液滴微流体技术对突变文库进行高通量筛选,在10h内,筛选了108个突变体,消耗了150μL反应试剂,与当时最先进的机器人筛选系统相比,筛选速度提高了1000倍,成本降低了100万倍,最终成功获得了催化效率提高10倍的优良突变体.这一成果充分展现了液滴微流体技术在高通量筛选领域的巨大的应用潜力.近年来,根据不同的筛选需要,研究人员对液滴微流体高通量筛选系统进行了一系列的优化改进.酶催化合成理想构型的手性化合物很大程度上依赖酶对手性异构体的选择性,但是天然具有较高选择性的催化酶很稀缺,大大限制了酶在催化手性药物合成中的应用.为了解决这一问题,Ma等[71]开发了一种双通道液滴微流体筛选系统(dual-channel microfluidic droplet screening,DMDS),用于筛选嗜热酯酶定向进化产生的突变文库,每天可筛选107个突变体,最终将嗜热酯酶对(S)-布洛芬底物的立体选择性提高了700倍.该系统使用蓝色和绿色双荧光底物作为报告信号,不同的信号输出,能够被双通道荧光检测系统高效分辨出来,进而分选出催化活性和立体选择性都显著提高的突变体. ...
Efficient molecular evolution to generate enantioselective enzymes using a dual-channel microfluidic droplet screening platform
1
2018
... 随着高效率封装单细胞技术的发展,单个细胞能被快速限制在微液滴内,实现细胞内代谢路径酶的高通量筛选.荧光底物被用来检测酶活性,其本身无荧光或者只有很低的荧光信号,但是其酶促反应产物则具有很强的荧光信号,根据反应前后荧光强度的变化,可以对酶活性进行分析和筛选.Agresti等[70]运用进化的方法对辣根过氧化物酶进行突变并转入酵母宿主体内,随后将荧光底物和酵母菌株一起封装在微液滴中,利用液滴微流体技术对突变文库进行高通量筛选,在10h内,筛选了108个突变体,消耗了150μL反应试剂,与当时最先进的机器人筛选系统相比,筛选速度提高了1000倍,成本降低了100万倍,最终成功获得了催化效率提高10倍的优良突变体.这一成果充分展现了液滴微流体技术在高通量筛选领域的巨大的应用潜力.近年来,根据不同的筛选需要,研究人员对液滴微流体高通量筛选系统进行了一系列的优化改进.酶催化合成理想构型的手性化合物很大程度上依赖酶对手性异构体的选择性,但是天然具有较高选择性的催化酶很稀缺,大大限制了酶在催化手性药物合成中的应用.为了解决这一问题,Ma等[71]开发了一种双通道液滴微流体筛选系统(dual-channel microfluidic droplet screening,DMDS),用于筛选嗜热酯酶定向进化产生的突变文库,每天可筛选107个突变体,最终将嗜热酯酶对(S)-布洛芬底物的立体选择性提高了700倍.该系统使用蓝色和绿色双荧光底物作为报告信号,不同的信号输出,能够被双通道荧光检测系统高效分辨出来,进而分选出催化活性和立体选择性都显著提高的突变体. ...
Microfluidic screening and whole-genome sequencing identifies mutations associated with improved protein secretion by yeast
1
2015
... 宏观尺度上代谢物的检测通常使用气质色谱或液质色谱这些大型仪器,虽然能够实现高精度、更广泛的代谢物检测,但是仪器的使用和维护成本非常高,而且样品制备耗时,筛选效率低下.液滴微流体技术的发展为代谢物的检测提供了新的途径,不仅可以实现更快的样品制备,而且液滴微反应器具有对单细胞进行分离区室化的功能,使不同突变的细胞可以在液滴微反应器中进行独立的培养,并且产生的代谢物在各自的微反应器中被隔开而不会相互干扰,因此在目标代谢物高产菌株的筛选中具有广阔的应用前景.如在对工业酶的生产菌株优化研究中,Sjostrom等[9]运用紫外诱变技术对表达α-淀粉酶的酵母菌株进行诱变,并且利用液滴微流体技术平台从含有105个突变体文库中筛选出一株α-淀粉酶表达量提高两倍的稳定高产菌株,该平台的筛选效率比自动微量滴定板筛选系统高出300倍,同时,筛选实验的试剂消耗量减少了1百万倍,大大降低了生产菌株筛选的成本.随后,Huang等[72]同样应用液滴微流体高通量筛选技术,经过多轮筛选,选择了8株α-淀粉酶分泌明显提高的菌株.通过对所有8株优良突变体进行全基因组测序,发现了330处基因突变,对突变的分析揭示了之前未曾发现的与蛋白分泌有关的生理机制,这将有助于研究人员设计更加高效的蛋白分泌细胞工厂,促进合成途径的理性构建.基于小分子代谢物生产和底物消耗对微生物菌株进行表型筛选是许多生物技术关键的瓶颈,Wang等[73]利用液滴微流体平台开发了一套高效筛选方法,用于对酿酒酵母木糖消耗和大肠杆菌乳酸生产的优良表型菌株的筛选.研究人员将荧光探针和单细胞菌株包裹在微反应器中,控制相同的孵育培养条件,通过对荧光信号的检测分析,能够评估微反应器里木糖或者乳酸浓度的变化,进而获得了理想表型的优良菌株. ...
Microfluidic high-throughput culturing of single cells for selection based on extracellular metabolite production or consumption
1
2014
... 宏观尺度上代谢物的检测通常使用气质色谱或液质色谱这些大型仪器,虽然能够实现高精度、更广泛的代谢物检测,但是仪器的使用和维护成本非常高,而且样品制备耗时,筛选效率低下.液滴微流体技术的发展为代谢物的检测提供了新的途径,不仅可以实现更快的样品制备,而且液滴微反应器具有对单细胞进行分离区室化的功能,使不同突变的细胞可以在液滴微反应器中进行独立的培养,并且产生的代谢物在各自的微反应器中被隔开而不会相互干扰,因此在目标代谢物高产菌株的筛选中具有广阔的应用前景.如在对工业酶的生产菌株优化研究中,Sjostrom等[9]运用紫外诱变技术对表达α-淀粉酶的酵母菌株进行诱变,并且利用液滴微流体技术平台从含有105个突变体文库中筛选出一株α-淀粉酶表达量提高两倍的稳定高产菌株,该平台的筛选效率比自动微量滴定板筛选系统高出300倍,同时,筛选实验的试剂消耗量减少了1百万倍,大大降低了生产菌株筛选的成本.随后,Huang等[72]同样应用液滴微流体高通量筛选技术,经过多轮筛选,选择了8株α-淀粉酶分泌明显提高的菌株.通过对所有8株优良突变体进行全基因组测序,发现了330处基因突变,对突变的分析揭示了之前未曾发现的与蛋白分泌有关的生理机制,这将有助于研究人员设计更加高效的蛋白分泌细胞工厂,促进合成途径的理性构建.基于小分子代谢物生产和底物消耗对微生物菌株进行表型筛选是许多生物技术关键的瓶颈,Wang等[73]利用液滴微流体平台开发了一套高效筛选方法,用于对酿酒酵母木糖消耗和大肠杆菌乳酸生产的优良表型菌株的筛选.研究人员将荧光探针和单细胞菌株包裹在微反应器中,控制相同的孵育培养条件,通过对荧光信号的检测分析,能够评估微反应器里木糖或者乳酸浓度的变化,进而获得了理想表型的优良菌株. ...
Single-cell multiomics: multiple measurements from single cells
1
2017
... 菌种进化工程利用各种化学和物理的诱变技术产生了大量的突变文库,为了从巨大的突变文库中获得有用的突变信息,寻找与优良表型相关联的分子机制,揭示单个细胞特有的属性,为进一步理性设计提供指导,需要高通量组学分选技术.由于液滴微流体技术具有快速的单细胞培养和捕获能力,可以将测序技术与液滴微流体系统结合,实现广泛的测序应用[74](如基因组、转录组、蛋白质组、表观遗传基因组),而且相比传统测序技术,在测序的通量、精确度和成本等方面也显现出巨大优势,极大促进了微生物组学分选领域的发展进步.Abate等[75]开发了一种基于液滴微流体技术的基因型分选系统,用于分析连续流动微液滴中的DNA分子.研究人员将荧光标记的DNA探针和目标模板共同封装在微液滴中.经历一系列反应后,如果探针与靶序列互补,则系统将检测到报告信号,随后对信号的振幅进行分析,能够分辨出碱基完全匹配或者错配的情况.通过扩展探针的组合,该系统可应用于对任意的未知目的序列的测序,从而为菌株基因型分选提供了快速且廉价的方法.此外,液滴微流体平台RNA测序也发展迅速,Treutlein等[76] 和Moon等[77]利用液滴微流体技术对细胞转录组进行测序,实现了对不同类型的细胞快速分选. ...
DNA sequence analysis with droplet-based microfluidics
1
2013
... 菌种进化工程利用各种化学和物理的诱变技术产生了大量的突变文库,为了从巨大的突变文库中获得有用的突变信息,寻找与优良表型相关联的分子机制,揭示单个细胞特有的属性,为进一步理性设计提供指导,需要高通量组学分选技术.由于液滴微流体技术具有快速的单细胞培养和捕获能力,可以将测序技术与液滴微流体系统结合,实现广泛的测序应用[74](如基因组、转录组、蛋白质组、表观遗传基因组),而且相比传统测序技术,在测序的通量、精确度和成本等方面也显现出巨大优势,极大促进了微生物组学分选领域的发展进步.Abate等[75]开发了一种基于液滴微流体技术的基因型分选系统,用于分析连续流动微液滴中的DNA分子.研究人员将荧光标记的DNA探针和目标模板共同封装在微液滴中.经历一系列反应后,如果探针与靶序列互补,则系统将检测到报告信号,随后对信号的振幅进行分析,能够分辨出碱基完全匹配或者错配的情况.通过扩展探针的组合,该系统可应用于对任意的未知目的序列的测序,从而为菌株基因型分选提供了快速且廉价的方法.此外,液滴微流体平台RNA测序也发展迅速,Treutlein等[76] 和Moon等[77]利用液滴微流体技术对细胞转录组进行测序,实现了对不同类型的细胞快速分选. ...
Reconstructing lineage hierarchies of the distal lung epithelium using single-cell RNA-seq
1
2014
... 菌种进化工程利用各种化学和物理的诱变技术产生了大量的突变文库,为了从巨大的突变文库中获得有用的突变信息,寻找与优良表型相关联的分子机制,揭示单个细胞特有的属性,为进一步理性设计提供指导,需要高通量组学分选技术.由于液滴微流体技术具有快速的单细胞培养和捕获能力,可以将测序技术与液滴微流体系统结合,实现广泛的测序应用[74](如基因组、转录组、蛋白质组、表观遗传基因组),而且相比传统测序技术,在测序的通量、精确度和成本等方面也显现出巨大优势,极大促进了微生物组学分选领域的发展进步.Abate等[75]开发了一种基于液滴微流体技术的基因型分选系统,用于分析连续流动微液滴中的DNA分子.研究人员将荧光标记的DNA探针和目标模板共同封装在微液滴中.经历一系列反应后,如果探针与靶序列互补,则系统将检测到报告信号,随后对信号的振幅进行分析,能够分辨出碱基完全匹配或者错配的情况.通过扩展探针的组合,该系统可应用于对任意的未知目的序列的测序,从而为菌株基因型分选提供了快速且廉价的方法.此外,液滴微流体平台RNA测序也发展迅速,Treutlein等[76] 和Moon等[77]利用液滴微流体技术对细胞转录组进行测序,实现了对不同类型的细胞快速分选. ...
Inertial-ordering-assisted droplet microfluidics for high-throughput single-cell RNA-sequencing
1
2018
... 菌种进化工程利用各种化学和物理的诱变技术产生了大量的突变文库,为了从巨大的突变文库中获得有用的突变信息,寻找与优良表型相关联的分子机制,揭示单个细胞特有的属性,为进一步理性设计提供指导,需要高通量组学分选技术.由于液滴微流体技术具有快速的单细胞培养和捕获能力,可以将测序技术与液滴微流体系统结合,实现广泛的测序应用[74](如基因组、转录组、蛋白质组、表观遗传基因组),而且相比传统测序技术,在测序的通量、精确度和成本等方面也显现出巨大优势,极大促进了微生物组学分选领域的发展进步.Abate等[75]开发了一种基于液滴微流体技术的基因型分选系统,用于分析连续流动微液滴中的DNA分子.研究人员将荧光标记的DNA探针和目标模板共同封装在微液滴中.经历一系列反应后,如果探针与靶序列互补,则系统将检测到报告信号,随后对信号的振幅进行分析,能够分辨出碱基完全匹配或者错配的情况.通过扩展探针的组合,该系统可应用于对任意的未知目的序列的测序,从而为菌株基因型分选提供了快速且廉价的方法.此外,液滴微流体平台RNA测序也发展迅速,Treutlein等[76] 和Moon等[77]利用液滴微流体技术对细胞转录组进行测序,实现了对不同类型的细胞快速分选. ...
Platforms for single-cell collection and analysis
1
2018
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
Single cell sequencing: a distinct new field
1
2017
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
Nanopore-based fourth-generation DNA sequencing technology
1
2015
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
Error rates for nanopore discrimination among cytosine, methylcytosine, and hydroxymethylcytosine along individual DNA strands
1
2013
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
Highly parallel direct RNA sequencing on an array of nanopores
1
2018
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
Analysis of RNA base modification and structural rearrangement by single-molecule real-time detection of reverse transcription
1
2013
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
Single-molecule site-specific detection of protein phosphorylation with a nanopore
1
2014
... 然而细胞某一方面的分子信息并不足以解释复杂的细胞行为,因为细胞状态是由基因组、表观基因组、转录组以及蛋白质组共同作用决定的,因此需要在多组学检测工具的集成和生物信息学数据的分析等方面取得更多地突破[78,79],为突变文库中异质性细胞绘制更全面的分子谱图,使我们对筛选出的细胞有更全面深入的分析.近些年,纳米孔道测序技术为多组学高通量集成检测提供了潜在的技术支撑,目前纳米孔道技术在DNA测序[80]和DNA修饰检测[81]、RNA测序[82]和RNA结构检测[83]、蛋白磷酸化检测[84]等方面有了许多突破性的进展,相信未来会极大地促进单细胞多组学检测技术在高通量筛选方面的应用. ...
eaaf4704
1
2017
... 随着合成生物学的发展,研究者对菌株的认识也越来越深入,尤其是近年来人工合成酵母的问世[85,86],可以让研究者更理性更高效地实现菌株的设计和大片段基因重组,产生更多样化的基因型和表型[87],这是今后在基础科学研究领域更好地研究基因结构变异和表型功能相互关系的重磅模版,并为菌株的快速进化带来了重大契机,但同时也给开发对应的高通量筛选平台提出了更高的挑战.虽然近年来高通量筛选方法的研究和应用都取得了很大的进展,但是一般只适用于特定的代谢物、酶或者代谢路径的筛选,缺乏普遍的适用性.未来随着生物信息学的快速发展,在生物传感器领域我们有更多的努力空间去尝试开发更多的特异性响应小分子的生物传感器,同时也应该在生物传感机制上做更多地探索,使得生物传感器的精度和准确度都能大幅提高.随着人工智能的发展以及人类对微生物体系越来越深入的认识,必将极大地推动自动化筛选和自动化数据分析能力的进步,为高通量筛选方法的建立提高有力的技术支撑. ...
eaaf4706
1
2017
... 随着合成生物学的发展,研究者对菌株的认识也越来越深入,尤其是近年来人工合成酵母的问世[85,86],可以让研究者更理性更高效地实现菌株的设计和大片段基因重组,产生更多样化的基因型和表型[87],这是今后在基础科学研究领域更好地研究基因结构变异和表型功能相互关系的重磅模版,并为菌株的快速进化带来了重大契机,但同时也给开发对应的高通量筛选平台提出了更高的挑战.虽然近年来高通量筛选方法的研究和应用都取得了很大的进展,但是一般只适用于特定的代谢物、酶或者代谢路径的筛选,缺乏普遍的适用性.未来随着生物信息学的快速发展,在生物传感器领域我们有更多的努力空间去尝试开发更多的特异性响应小分子的生物传感器,同时也应该在生物传感机制上做更多地探索,使得生物传感器的精度和准确度都能大幅提高.随着人工智能的发展以及人类对微生物体系越来越深入的认识,必将极大地推动自动化筛选和自动化数据分析能力的进步,为高通量筛选方法的建立提高有力的技术支撑. ...
Precise control of SCRaMbLE in synthetic haploid and diploid yeast
1
2018
... 随着合成生物学的发展,研究者对菌株的认识也越来越深入,尤其是近年来人工合成酵母的问世[85,86],可以让研究者更理性更高效地实现菌株的设计和大片段基因重组,产生更多样化的基因型和表型[87],这是今后在基础科学研究领域更好地研究基因结构变异和表型功能相互关系的重磅模版,并为菌株的快速进化带来了重大契机,但同时也给开发对应的高通量筛选平台提出了更高的挑战.虽然近年来高通量筛选方法的研究和应用都取得了很大的进展,但是一般只适用于特定的代谢物、酶或者代谢路径的筛选,缺乏普遍的适用性.未来随着生物信息学的快速发展,在生物传感器领域我们有更多的努力空间去尝试开发更多的特异性响应小分子的生物传感器,同时也应该在生物传感机制上做更多地探索,使得生物传感器的精度和准确度都能大幅提高.随着人工智能的发展以及人类对微生物体系越来越深入的认识,必将极大地推动自动化筛选和自动化数据分析能力的进步,为高通量筛选方法的建立提高有力的技术支撑. ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}